Evolution
and the Origins of Disease
The principles of
evolution by natural selection are finally beginning to inform medicine by
Randolph M. Nesse and George C. Williams ...........

Thoughtful
contemplation of the human body elicits awe--in equal measure with perplexity.
The eye, for instance, has long been an object of wonder, with the clear,
living tissue of the cornea curving just the right amount, the iris adjusting
to brightness and the lens to distance, so that the optimal quantity of light focuses
exactly on the surface of the retina. Admiration of such apparent perfection
soon gives way, however, to consternation. Contrary to any sensible design,
blood vessels and nerves traverse the inside of the retina, creating a blind
spot at their point of exit.
The body is a
bundle of such jarring contradictions. For each exquisite heart valve, we have
a wisdom tooth. Strands of DNA direct the development of the 10 trillion cells
that make up a human adult but then permit his or her steady deterioration and
eventual death. Our immune system can identify and destroy a million kinds of
foreign matter, yet many bacteria can still kill us. These contradictions make
it appear as if the body was designed by a team of superb engineers with
occasional interventions by Rube Goldberg.
In fact, such
seeming incongruities make sense but only when we investigate the origins of
the body's vulnerabilities while keeping in mind the wise words of
distinguished geneticist Theodosius Dobzhansky: "Nothing in biology makes sense
except in the light of evolution." Evolutionary biology is, of course, the
scientific foundation for all biology, and biology is the foundation for all
medicine. To a surprising degree, however, evolutionary biology is just now
being recognized as a basic medical science. The enterprise of studying medical
problems in an evolutionary context has been termed Darwinian medicine. Most
medical research tries to explain the causes of an individual's disease and
seeks therapies to cure or relieve deleterious conditions. These efforts are
traditionally based on consideration of proximate issues, the straightforward
study of the body's anatomic and physiological mechanisms as they currently
exist. In contrast, Darwinian medicine asks why the body is designed in a way
that makes us all vulnerable to problems like cancer, atherosclerosis,
depression and choking, thus offering a broader context in which to conduct
research.

DEFENSIVE
RESPONSES
The evolutionary
explanations for the body's flaws fall into surprisingly few categories. First,
some discomforting conditions, such as pain, fever, cough, vomiting and
anxiety, are actually neither diseases nor design defects but rather are
evolved defenses. Second, conflicts with other organisms--Escherichia coli or crocodiles,
for instance--are a fact of life. Third, some circumstances, such as the ready
availability of dietary fats, are so recent that natural selection has not yet
had a chance to deal with them. Fourth, the body may fall victim to trade-offs
between a trait's benefits and its costs; a textbook example is the sickle cell
gene, which also protects against malaria. Finally, the process of natural
selection is constrained in ways that leave us with suboptimal design features,
as in the case of the mammalian eye.
Evolved Defenses
Perhaps the most
obviously useful defense mechanism is coughing; people who cannot clear foreign
matter from their lungs are likely to die from pneumonia. The capacity for pain
is also certainly beneficial. The rare individuals who cannot feel pain fail
even to experience discomfort from staying in the same position for long
periods. Their unnatural stillness impairs the blood supply to their joints,
which then deteriorate. Such pain-free people usually die by early adulthood from
tissue damage and infections. Cough or pain is usually interpreted as disease
or trauma but is actually part of the solution rather than the problem. These
defensive capabilities, shaped by natural selection, are kept in reserve until
needed.
Less widely
recognized as defenses are fever, nausea, vomiting, diarrhea, anxiety, fatigue,
sneezing and inflammation. Even some physicians remain unaware of fever's
utility. No mere increase in metabolic rate, fever is a carefully regulated
rise in the set point of the body's thermostat. The higher body temperature
facilitates the destruction of pathogens. Work by Matthew J. Kluger of the
Lovelace Institute in Albuquerque, N.M., has shown that even cold-blooded
lizards, when infected, move to warmer places until their bodies are several
degrees above their usual temperature. If prevented from moving to the warm
part of their cage, they are at increased risk of death from the infection. In
a similar study by Evelyn Satinoff of the University of Delaware, elderly rats,
who can no longer achieve the high fevers of their younger lab companions, also
instinctively sought hotter environments when challenged by infection.
A reduced level
of iron in the blood is another misunderstood defense mechanism. People
suffering from chronic infection often have decreased levels of blood iron.
Although such low iron is sometimes blamed for the illness, it actually is a
protective response: during infection, iron is sequestered in the liver, which
prevents invading bacteria from getting adequate supplies of this vital
element.
Morning sickness
has long been considered an unfortunate side effect of pregnancy. The nausea,
however, coincides with the period of rapid tissue differentiation of the
fetus, when development is most vulnerable to interference by toxins. And
nauseated women tend to restrict their intake of strong-tasting, potentially
harmful substances. These observations led independent researcher Margie Profet
to hypothesize that the nausea of pregnancy is an adaptation whereby the mother
protects the fetus from exposure to toxins. Profet tested this idea by
examining pregnancy outcomes. Sure enough, women with more nausea were less
likely to suffer miscarriages. (This evidence supports the hypothesis but is
hardly conclusive. If Profet is correct, further research should discover that
pregnant females of many species show changes in food preferences. Her theory
also predicts an increase in birth defects among offspring of women who have
little or no morning sickness and thus eat a wider variety of foods during
pregnancy.)
Another common
condition, anxiety, obviously originated as a defense in dangerous situations
by promoting escape and avoidance. A 1992 study by Lee A. Dugatkin of the
University of Louisville evaluated the benefits of fear in guppies. He grouped
them as timid, ordinary or bold, depending on their reaction to the presence of
smallmouth bass. The timid hid, the ordinary simply swam away, and the bold
maintained their ground and eyed the bass. Each guppy group was then left alone
in a tank with a bass. After 60 hours, 40 percent of the timid guppies had
survived, as had only 15 percent of the ordinary fish. The entire complement of
bold guppies, on the other hand, wound up aiding the transmission of bass genes
rather than their own.
Selection for
genes promoting anxious behaviors implies that there should be people who
experience too much anxiety, and indeed there are. There should also be
hypophobic individuals who have insufficient anxiety, either because of genetic
tendencies or antianxiety drugs. The exact nature and frequency of such a
syndrome is an open question, as few people come to psychiatrists complaining
of insufficient apprehension. But if sought, the pathologically nonanxious may
be found in emergency rooms, jails and unemployment lines.
The utility of
common and unpleasant conditions such as diarrhea, fever and anxiety is not
intuitive. If natural selection shapes the mechanisms that regulate defensive
responses, how can people get away with using drugs to block these defenses
without doing their bodies obvious harm? Part of the answer is that we do, in
fact, sometimes do ourselves a disservice by disrupting defenses.
Herbert L. DuPont
of the University of Texas at Houston and Richard B. Hornick of Orlando
Regional Medical Center studied the diarrhea caused by Shigella infection and
found that people who took antidiarrhea drugs stayed sick longer and were more
likely to have complications than those who took a placebo. In another example,
Eugene D. Weinberg of Indiana University has documented that well-intentioned
attempts to correct perceived iron deficiencies have led to increases in
infectious disease, especially amebiasis, in parts of Africa. Although the iron
in most oral supplements is unlikely to make much difference in otherwise
healthy people with everyday infections, it can severely harm those who are
infected and malnourished. Such people cannot make enough protein to bind the
iron, leaving it free for use by infectious agents.
On the
morning-sickness front, an antinausea drug was recently blamed for birth
defects. It appears that no consideration was given to the possibility that the
drug itself might be harmless to the fetus but could still be associated with
birth defects, by interfering with the mother's defensive nausea.
Another obstacle
to perceiving the benefits of defenses arises from the observation that many
individuals regularly experience seemingly worthless reactions of anxiety,
pain, fever, diarrhea or nausea. The explanation requires an analysis of the
regulation of defensive responses in terms of signal-detection theory. A
circulating toxin may come from something in the stomach. An organism can expel
it by vomiting, but only at a price. The cost of a false alarm--vomiting when
no toxin is truly present--is only a few calories. But the penalty for a single
missed authentic alarm--failure to vomit when confronted with a toxin--may be
death.
Natural selection
therefore tends to shape regulation mechanisms with hair triggers, following
what we call the smoke-detector principle. A smoke alarm that will reliably
wake a sleeping family in the event of any fire will necessarily give a false
alarm every time the toast burns. The price of the human body's numerous
"smoke alarms" is much suffering that is completely normal but in
most instances unnecessary. This principle also explains why blocking defenses
is so often free of tragic consequences. Because most defensive reactions occur
in response to insignificant threats, interference is usually harmless; the
vast majority of alarms that are stopped by removing the battery from the smoke
alarm are false ones, so this strategy may seem reasonable. Until, that is, a
real fire occurs.
Conflicts with
Other Organisms
Natural selection
is unable to provide us with perfect protection against all pathogens, because
they tend to evolve much faster than humans do. E. coli, for example, with its
rapid rates of reproduction, has as much opportunity for mutation and selection
in one day as humanity gets in a millennium. And our defenses, whether natural
or artificial, make for potent selection forces. Pathogens either quickly
evolve a counterdefense or become extinct. Amherst College biologist Paul W.
Ewald has suggested classifying phenomena associated with infection according
to whether they benefit the host, the pathogen, both or neither. Consider the
runny nose associated with a cold. Nasal mucous secretion could expel
intruders, speed the pathogen's transmission to new hosts or both [see
"The Evolution of Virulence," by Paul W. Ewald; Scientific American,
April 1993]. Answers could come from studies examining whether blocking nasal
secretions shortens or prolongs illness, but few such studies have been done.
EVOLUTION
OF VIRULENCE

Humanity won huge
battles in the war against pathogens with the development of antibiotics and
vaccines. Our victories were so rapid and seemingly complete that in 1969 U.S.
Surgeon General William H. Stewart said that it was "time to close the
book on infectious disease." But the enemy, and the power of natural
selection, had been underestimated. The sober reality is that pathogens
apparently can adapt to every chemical researchers develop. ("The war has
been won," one scientist more recently quipped. "By the other
side.")
Antibiotic
resistance is a classic demonstration of natural selection. Bacteria that
happen to have genes that allow them to prosper despite the presence of an
antibiotic reproduce faster than others, and so the genes that confer resistance
spread quickly. As shown by Nobel laureate Joshua Lederberg of the Rockefeller
University, they can even jump to different species of bacteria, borne on bits
of infectious DNA. Today some strains of tuberculosis in New York City are
resistant to all three main antibiotic treatments; patients with those strains
have no better chance of surviving than did TB patients a century ago. Stephen
S. Morse of Columbia University notes that the multidrug-resistant strain that
has spread throughout the East Coast may have originated in a homeless shelter
across the street from Columbia-Presbyterian Medical Center. Such a phenomenon
would indeed be predicted in an environment where fierce selection pressure
quickly weeds out less hardy strains. The surviving bacilli have been bred for
resistance.
Many people,
including some physicians and scientists, still believe the outdated theory
that pathogens necessarily become benign after long association with hosts.
Superficially, this makes sense. An organism that kills rapidly may never get
to a new host, so natural selection would seem to favor lower virulence.
Syphilis, for instance, was a highly virulent disease when it first arrived in
Europe, but as the centuries passed it became steadily more mild. The virulence
of a pathogen is, however, a life history trait that can increase as well as
decrease, depending on which option is more advantageous to its genes.
For agents of
disease that are spread directly from person to person, low virulence tends to
be beneficial, as it allows the host to remain active and in contact with other
potential hosts. But some diseases, like malaria, are transmitted just as
well--or better--by the incapacitated. For such pathogens, which usually rely
on intermediate vectors like mosquitoes, high virulence can give a selective
advantage. This principle has direct implications for infection control in
hospitals, where health care workers' hands can be vectors that lead to
selection for more virulent strains.
In the case of
cholera, public water supplies play the mosquitoes' role. When water for
drinking and bathing is contaminated by waste from immobilized patients,
selection tends to increase virulence, because more diarrhea enhances the
spread of the organism even if individual hosts quickly die. But, as Ewald has
shown, when sanitation improves, selection acts against classical Vibrio
cholerae bacteria in favor of the more benign El Tor biotype. Under these
conditions, a dead host is a dead end. But a less ill and more mobile host,
able to infect many others over a much longer time, is an effective vehicle for
a pathogen of lower virulence. In another example, better sanitation leads to
displacement of the aggressive Shigella flexneri by the more benign S. sonnei.
NEW
ENVIRONMENTS, NEW THREATS
Such
considerations may be relevant for public policy. Evolutionary theory predicts
that clean needles and the encouragement of safe sex will do more than save
numerous individuals from HIV infection. If humanity's behavior itself slows
HIV transmission rates, strains that do not soon kill their hosts have the
long-term survival advantage over the more virulent viruses that then die with
their hosts, denied the opportunity to spread. Our collective choices can
change the very nature of HIV.
Conflicts with
other organisms are not limited to pathogens. In times past, humans were at
great risk from predators looking for a meal. Except in a few places, large
carnivores now pose no threat to humans. People are in more danger today from
smaller organisms' defenses, such as the venoms of spiders and snakes.
Ironically, our fears of small creatures, in the form of phobias, probably
cause more harm than any interactions with those organisms do. Far more
dangerous than predators or poisoners are other members of our own species. We
attack each other not to get meat but to get mates, territory and other
resources. Violent conflicts between individuals are overwhelmingly between
young men in competition and give rise to organizations to advance these aims.
Armies, again usually composed of young men, serve similar objectives, at huge
cost.
Even the most
intimate human relationships give rise to conflicts having medical
implications. The reproductive interests of a mother and her infant, for
instance, may seem congruent at first but soon diverge. As noted by biologist
Robert L. Trivers in a now classic 1974 paper, when her child is a few years
old, the mother's genetic interests may be best served by becoming pregnant
again, whereas her offspring benefits from continuing to nurse. Even in the
womb there is contention. From the mother's vantage point, the optimal size of
a fetus is a bit smaller than that which would best serve the fetus and the
father. This discord, according to David Haig of Harvard University, gives rise
to an arms race between fetus and mother over her levels of blood pressure and
blood sugar, sometimes resulting in hypertension and diabetes during pregnancy.
Coping with
Novelty
Making rounds in
any modern hospital provides sad testimony to the prevalence of diseases
humanity has brought on itself. Heart attacks, for example, result mainly from
atherosclerosis, a problem that became widespread only in this century and that
remains rare among hunter-gatherers. Epidemiological research furnishes the
information that should help us prevent heart attacks: limit fat intake, eat
lots of vegetables, and exercise hard each day. But hamburger chains
proliferate, diet foods languish on the shelves, and exercise machines serve as
expensive clothing hangers throughout the land. The proportion of overweight
Americans is one third and rising. We all know what is good for us. Why do so
many of us continue to make unhealthy choices?
Our poor
decisions about diet and exercise are made by brains shaped to cope with an
environment substantially different from the one our species now inhabits. On
the African savanna, where the modern human design was fine-tuned, fat, salt
and sugar were scarce and precious. Individuals who had a tendency to consume
large amounts of fat when given the rare opportunity had a selective advantage.
They were more likely to survive famines that killed their thinner companions.
And we, their descendants, still carry those urges for foodstuffs that today
are anything but scarce. These evolved desires--inflamed by advertisements from
competing food corporations that themselves survive by selling us more of
whatever we want to buy--easily defeat our intellect and willpower. How ironic
that humanity worked for centuries to create environments that are almost
literally flowing with milk and honey, only to see our success responsible for
much modern disease and untimely death.
Increasingly,
people also have easy access to many kinds of drugs, especially alcohol and
tobacco, that are responsible for a huge proportion of disease, health care
costs and premature death. Although individuals have always used psychoactive
substances, widespread problems materialized only following another
environmental novelty: the ready availability of concentrated drugs and new,
direct routes of administration, especially injection. Most of these
substances, including nicotine, cocaine and opium, are products of natural
selection that evolved to protect plants from insects. Because humans share a
common evolutionary heritage with insects, many of these substances also affect
our nervous system.
This perspective
suggests that it is not just defective individuals or disordered societies that
are vulnerable to the dangers of psychoactive drugs; all of us are susceptible
because drugs and our biochemistry have a long history of interaction.
Understanding the details of that interaction, which is the focus of much
current research from both a proximate and evolutionary perspective, may well
lead to better treatments for addiction.
The relatively
recent and rapid increase in breast cancer must be the result in large part of
changing environments and ways of life, with only a few cases resulting solely
from genetic abnormalities. Boyd Eaton and his colleagues at Emory University
reported that the rate of breast cancer in today's "nonmodern"
societies is only a tiny fraction of that in the U.S. They hypothesize that the
amount of time between menarche and first pregnancy is a crucial risk factor,
as is the related issue of total lifetime number of menstrual cycles. In
hunter-gatherers, menarche occurs at about age 15 or later, followed within a
few years by pregnancy and two or three years of nursing, then by another
pregnancy soon after. Only between the end of nursing and the next pregnancy
will the woman menstruate and thus experience the high levels of hormones that
may adversely affect breast cells.
In modern
societies, in contrast, menarche occurs at age 12 or 13--probably at least in
part because of a fat intake sufficient to allow an extremely young woman to
nourish a fetus--and the first pregnancy may be decades later or never. A
female hunter-gatherer may have a total of 150 menstrual cycles, whereas the
average woman in modern societies has 400 or more. Although few would suggest
that women should become pregnant in their teens to prevent breast cancer
later, early administration of a burst of hormones to simulate pregnancy may
reduce the risk. Trials to test this idea are now under way at the University
of California at San Diego.
Trade-offs and
Constraints
Compromise is
inherent in every adaptation. Arm bones three times their current thickness
would almost never break, but Homo sapiens would be lumbering creatures on a
never-ending quest for calcium. More sensitive ears might sometimes be useful,
but we would be distracted by the noise of air molecules banging into our
eardrums.
Such trade-offs
also exist at the genetic level. If a mutation offers a net reproductive
advantage, it will tend to increase in frequency in a population even if it
causes vulnerability to disease. People with two copies of the sickle cell
gene, for example, suffer terrible pain and die young. People with two copies
of the "normal" gene are at high risk of death from malaria. But individuals
with one of each are protected from both malaria and sickle cell disease. Where
malaria is prevalent, such people are fitter, in the Darwinian sense, than
members of either other group. So even though the sickle cell gene causes
disease, it is selected for where malaria persists. Which is the
"healthy" allele in this environment? The question has no answer.
There is no one normal human genome--there are only genes.
SMALL
APPENDIX
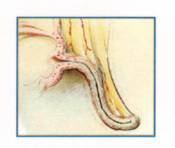
Many other genes
that cause disease must also have offered benefits, at least in some
environments, or they would not be so common. Because cystic fibrosis (CF)
kills one out of 2,500 Caucasians, the responsible genes would appear to be at
great risk of being eliminated from the gene pool. And yet they endure. For years,
researchers mused that the CF gene, like the sickle cell gene, probably
conferred some advantage. Recently a study by Gerald B. Pier of Harvard Medical
School and his colleagues gave substance to this informed speculation: having
one copy of the CF gene appears to decrease the chances of the bearer acquiring
a typhoid fever infection, which once had a 15 percent mortality.
Aging may be the
ultimate example of a genetic trade-off. In 1957 one of us (Williams) suggested
that genes that cause aging and eventual death could nonetheless be selected
for if they had other effects that gave an advantage in youth, when the force
of selection is stronger. For instance, a hypothetical gene that governs
calcium metabolism so that bones heal quickly but that also happens to cause
the steady deposition of calcium in arterial walls might well be selected for
even though it kills some older people. The influence of such pleiotropic genes
(those having multiple effects) has been seen in fruit flies and flour beetles,
but no specific example has yet been found in humans. Gout, however, is of
particular interest, because it arises when a potent antioxidant, uric acid,
forms crystals that precipitate out of fluid in joints. Antioxidants have
antiaging effects, and plasma levels of uric acid in different species of
primates are closely correlated with average adult life span. Perhaps high
levels of uric acid benefit most humans by slowing tissue aging, while a few
pay the price with gout.
Other examples
are more likely to contribute to more rapid aging. For instance, strong immune
defenses protect us from infection but also inflict continuous, low-level
tissue damage. It is also possible, of course, that most genes that cause aging
have no benefit at any age--they simply never decreased reproductive fitness
enough in the natural environment to be selected against. Nevertheless, over
the next decade research will surely identify specific genes that accelerate
senescence, and researchers will soon thereafter gain the means to interfere
with their actions or even change them. Before we tinker, however, we should
determine whether these actions have benefits early in life.
Because evolution
can take place only in the direction of time's arrow, an organism's design is
constrained by structures already in place. As noted, the vertebrate eye is
arranged backward. The squid eye, in contrast, is free from this defect, with
vessels and nerves running on the outside, penetrating where necessary and
pinning down the retina so it cannot detach. The human eye's flaw results from
simple bad luck; hundreds of millions of years ago, the layer of cells that
happened to become sensitive to light in our ancestors was positioned
differently from the corresponding layer in ancestors of squids. The two
designs evolved along separate tracks, and there is no going back.
Such path
dependence also explains why the simple act of swallowing can be
life-threatening. Our respiratory and food passages intersect because in an
early lungfish ancestor the air opening for breathing at the surface was
understandably located at the top of the snout and led into a common space
shared by the food passageway. Because natural selection cannot start from
scratch, humans are stuck with the possibility that food will clog the opening
to our lungs.
The path of
natural selection can even lead to a potentially fatal cul-de-sac, as in the
case of the appendix, that vestige of a cavity that our ancestors employed in
digestion. Because it no longer performs that function, and as it can kill when
infected, the expectation might be that natural selection would have eliminated
it. The reality is more complex. Appendicitis results when inflammation causes
swelling, which compresses the artery supplying blood to the appendix. Blood
flow protects against bacterial growth, so any reduction aids infection, which
creates more swelling. If the blood supply is cut off completely, bacteria have
free rein until the appendix bursts. A slender appendix is especially
susceptible to this chain of events, so appendicitis may, paradoxically, apply
the selective pressure that maintains a large appendix. Far from arguing that
everything in the body is perfect, an evolutionary analysis reveals that we
live with some very unfortunate legacies and that some vulnerabilities may even
be actively maintained by the force of natural selection.
Evolution of
Darwinian Medicine
Despite the power
of the Darwinian paradigm, evolutionary biology is just now being recognized as
a basic science essential for medicine. Most diseases decrease fitness, so it
would seem that natural selection could explain only health, not disease. A
Darwinian approach makes sense only when the object of explanation is changed
from diseases to the traits that make us vulnerable to diseases. The assumption
that natural selection maximizes health also is incorrect--selection maximizes
the reproductive success of genes. Those genes that make bodies having superior
reproductive success will become more common, even if they compromise the
individual's health in the end.
Finally, history
and misunderstanding have presented obstacles to the acceptance of Darwinian
medicine. An evolutionary approach to functional analysis can appear akin to
naive teleology or vitalism, errors banished only recently, and with great
effort, from medical thinking. And, of course, whenever evolution and medicine
are mentioned together, the specter of eugenics arises. Discoveries made
through a Darwinian view of how all human bodies are alike in their vulnerability
to disease will offer great benefits for individuals, but such insights do not
imply that we can or should make any attempt to improve the species. If
anything, this approach cautions that apparent genetic defects may have
unrecognized adaptive significance, that a single "normal" genome is
nonexistent and that notions of "normality" tend to be simplistic.
The systematic
application of evolutionary biology to medicine is a new enterprise. Like
biochemistry at the beginning of this century, Darwinian medicine very likely
will need to develop in several incubators before it can prove its power and
utility. If it must progress only from the work of scholars without funding to
gather data to test their ideas, it will take decades for the field to mature.
Departments of evolutionary biology in medical schools would accelerate the
process, but for the most part they do not yet exist. If funding agencies had
review panels with evolutionary expertise, research would develop faster, but
such panels remain to be created. We expect that they will.
The evolutionary
viewpoint provides a deep connection between the states of disease and normal
functioning and can integrate disparate avenues of medical research as well as
suggest fresh and important areas of inquiry. Its utility and power will
ultimately lead to recognition of evolutionary biology as a basic medical
science.
The Authors
RANDOLPH M. NESSE
and GEORGE C. WILLIAMS are the authors of the 1994 book Why We Get Sick: The
New Science of Darwinian Medicine. Nesse received his medical degree from the
University of Michigan Medical School in 1974. He is now professor of
psychiatry at that institution and is director of the Evolution and Human
Adaptation Program at the university's Institute for Social Research. Williams
received his doctorate in 1955 from the University of California, Los Angeles,
and quickly became one of the world's foremost evolutionary theorists. A member
of the National Academy of Sciences, he is professor emeritus of ecology and
evolution at the State University of New York at Stony Brook and edits the
Quarterly Review of Biology.